The synaptic connections that define such circuits are typically made in a dense tangle of dendrites, axons terminals, and glial cell processes that together constitute what is called neuropil.
Knee-Jerk Response
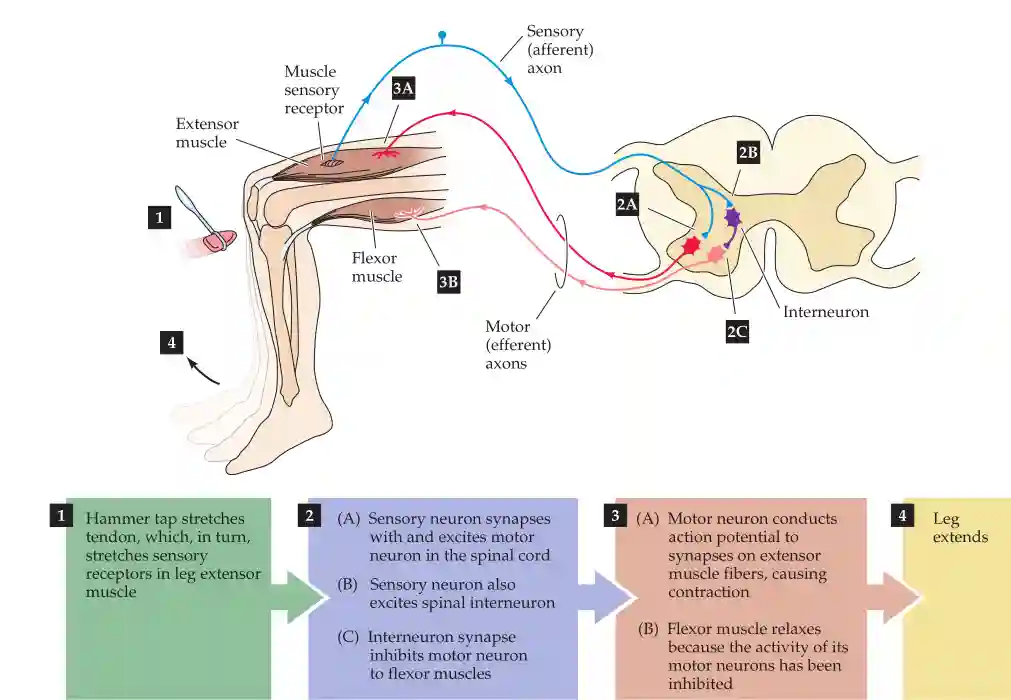
The knee-jerk reflex (also known as the patellar reflex) is a classic example of a mono-synaptic reflex arc, which involves a direct connection between sensory and motor neurons, as well as inhibitory circuits to regulate movement. This is important for us if we fall and hit something.
Neural Circuit of the Knee-Jerk Reflex
The reflex involves the following components:
- Sensory Neuron (Afferent Pathway)
- When the patellar tendon is tapped, muscle spindles in the quadriceps detect the sudden stretch.
- This activates Ia afferent fibers, which send an electrical signal to the spinal cord.
- Interneuron (Inhibitory Pathway)
- Within the spinal cord, the afferent neuron synapses onto an inhibitory interneuron.
- This interneuron, in turn, inhibits the motor neuron controlling the antagonist muscle (flexor muscle, hamstring) to prevent opposing contraction.
- Motor Neuron (Efferent Pathway to Extensor Muscle)
- The direct monosynaptic connection between the sensory neuron and the alpha motor neuron in the spinal cord leads to activation of the quadriceps muscle.
- This results in contraction of the extensor muscle (quadriceps), producing the characteristic leg kick.
- Motor Neuron (Efferent Pathway to Flexor Muscle - Inhibition)
- The inhibitory interneuron prevents contraction of the antagonist muscle (hamstring), ensuring smooth movement.
This reflex demonstrates the fundamental organization of afferent (sensory) and efferent (motor) pathways, where information flows toward the central nervous system (CNS) via afferents and away from it via efferents.
The inhibitory interneuron releases neurotransmitters like GABA or glycine, which cause hyperpolarization of the flexor motor neuron.
- This inhibits the hamstring contraction, allowing smooth knee extension.
- The hyperpolarization prevents unwanted co-contraction of opposing muscles.
Recording Methods for Neural Activity
Neural activity can be recorded during reflex responses, including in clinical and surgical settings.
There are two primary methods:
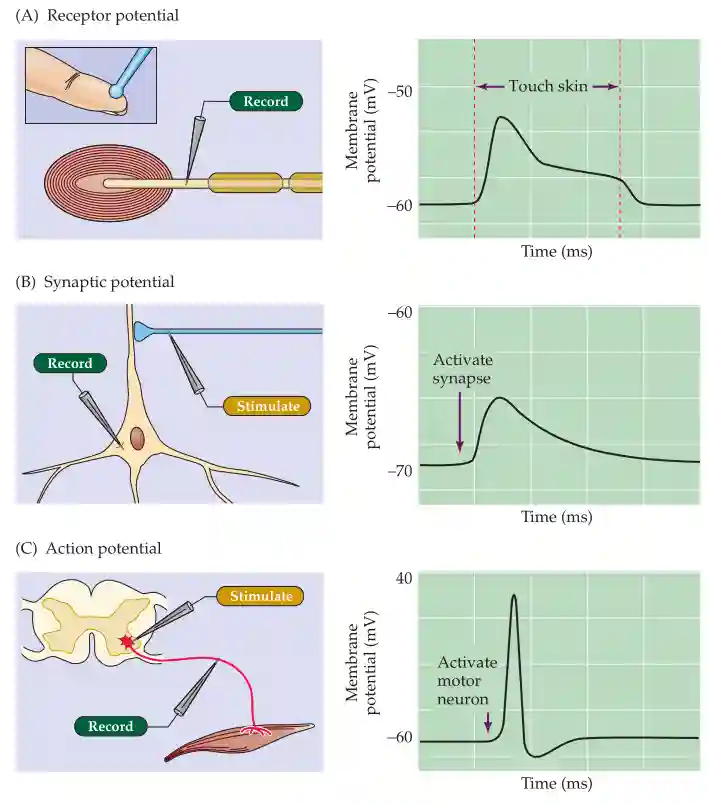
- Extracellular Recording with Microelectrodes
- A glass micro-electrode with a super-thin metal tip is used to measure the voltage difference between the inside and outside of a neuron.
- This is mainly used to record action potentials or field potentials from neurons.
- Intracellular Current Injection
- A current can be injected into the neuron to measure its passive properties (such as resistance and capacitance) and active properties (such as action potential threshold and firing patterns).
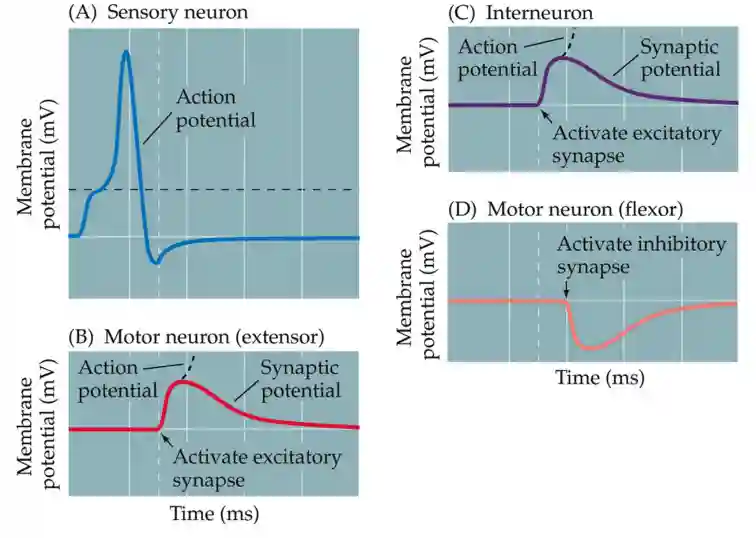
Intracellular recordings can detect the smaller, graded potential changes that trigger action potentials, and thus allow a more detailed analy- sis of communication between neurons within a circuit.
In a similar way, we measure the myotatic circuits in the heart.
These techniques have also been applied in the human brain during surgery, such as in procedures for epilepsy treatment or deep brain stimulation, where patients can report their sensations in real time while neuronal activity is recorded.
Activations in the Knee-Jerk response
Receptive Fields
The receptive field of a neuron refers to the specific region of the body (e.g., skin, retina) that, when stimulated, makes the neuron fire.
For example:
- In the somatosensory system, a mechanoreceptor in the skin will fire when a specific area is touched.
- In the visual system, a retinal ganglion cell has a receptive field defined by a portion of the visual space.
Key properties of receptive fields:
- Size and resolution: Smaller receptive fields (e.g., in fingertips) allow higher spatial resolution and finer touch discrimination.
- Excitatory and inhibitory zones: Many neurons have center-surround organization, where stimulation of the center excites the neuron, while stimulation of the surrounding area inhibits it.
The single neuron electrophysiological recording is opposed to functional brain imaging.
Electrical Signals in Neurons
Neurons communicate through electrical signals that arise due to the movement of ions across the cell membrane. This is achieved through:
Types of potentials and signals
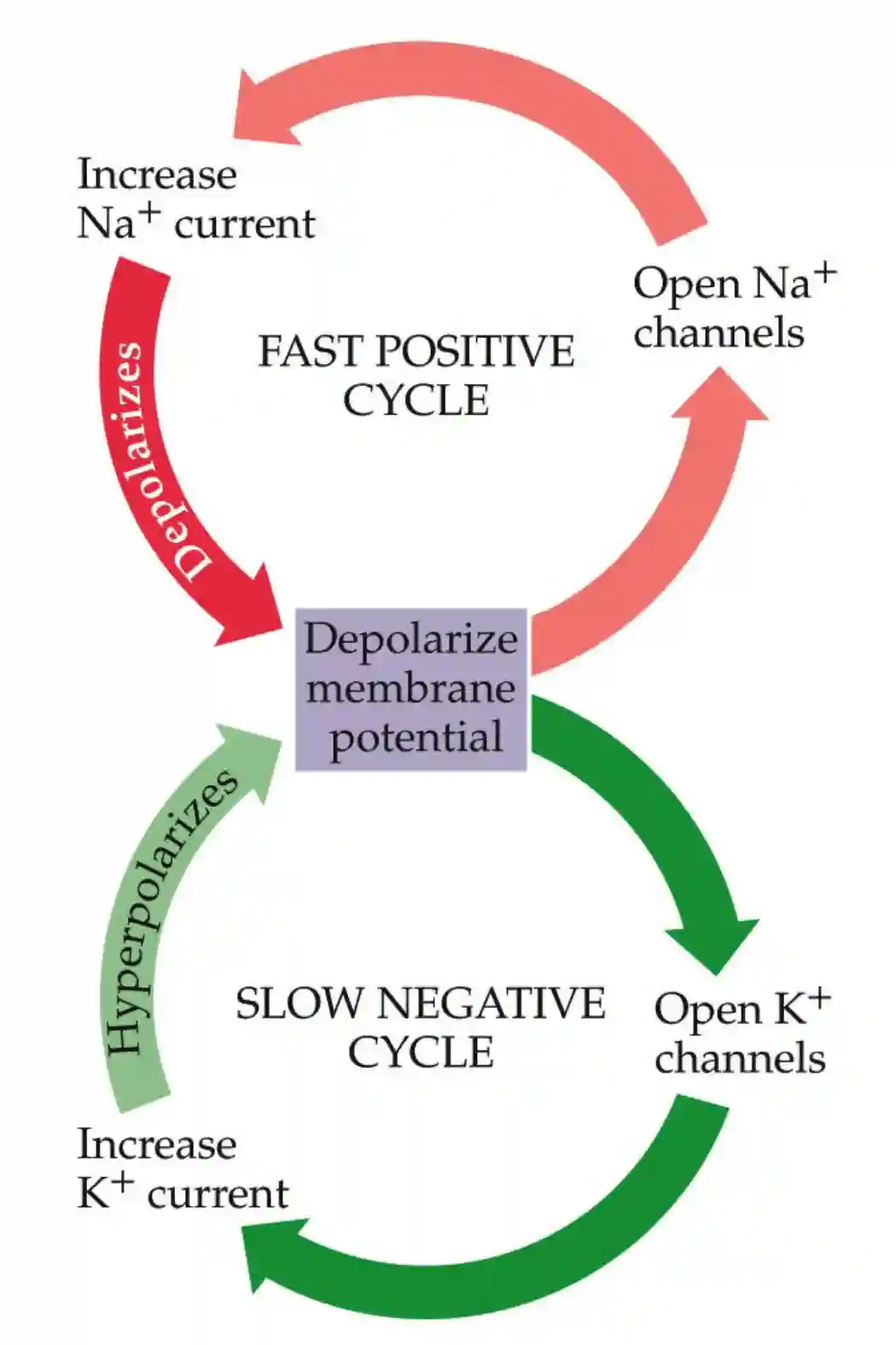
In general, action potentials are divided in two phases, a first Na phase and second potassium phase.
Potentials
- Receptor Potentials (input signals from sensory receptors)
- Synaptic Potentials (signals from other neurons)
- Action Potentials (output signals transmitted down the axon)
A fundamental problem for neurons is that their axons, which can be quite long (remember that a spinal motor neuron can extend for a meter or more), are not good electrical conductors. [...] To compensate for this deficiency, neurons have evolved a “booster system” that allows them to conduct electrical signals over great distances despite their intrinsically poor electrical characteristics. he electrical signals produced by this booster system are called action potentials
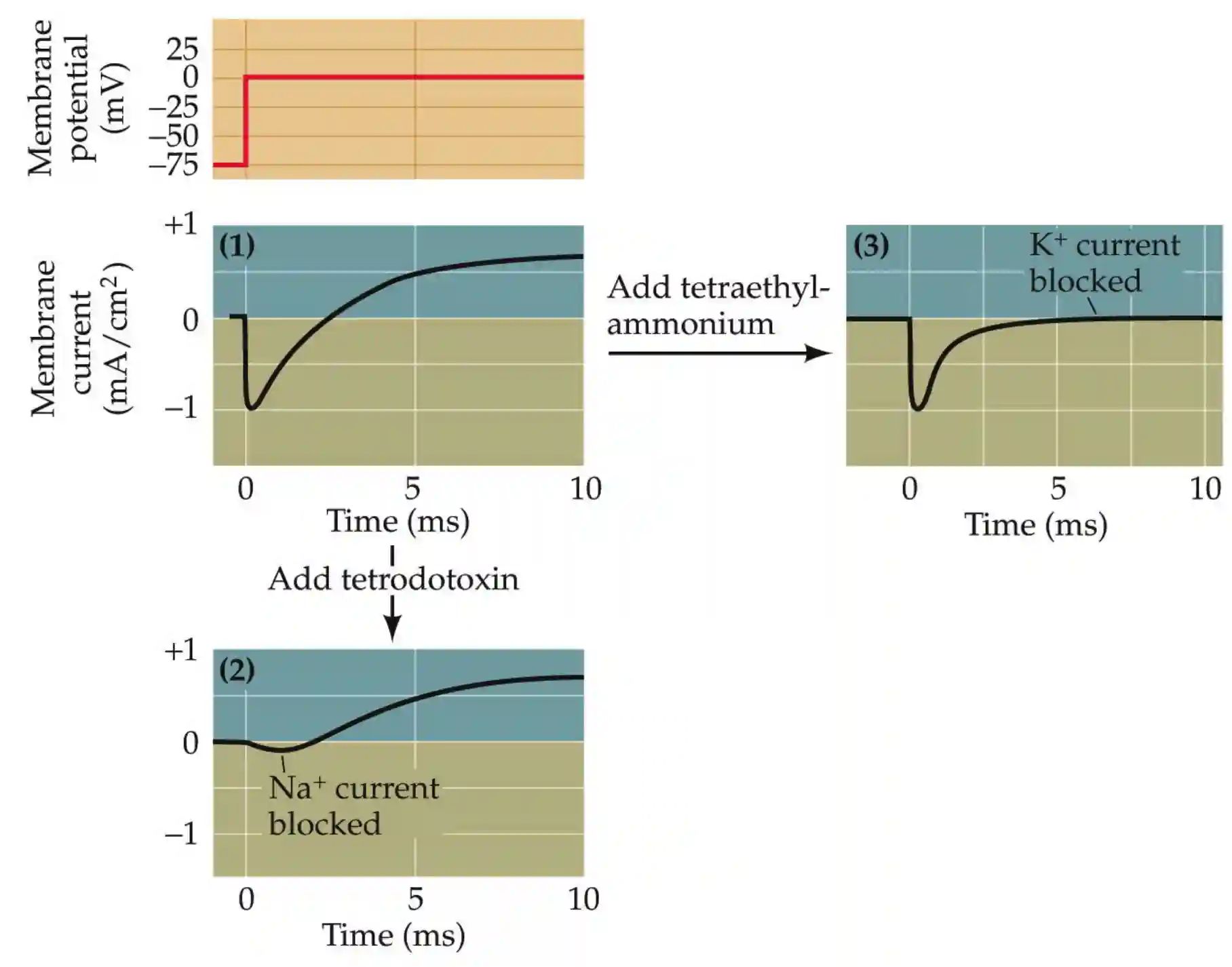
Role of sodium
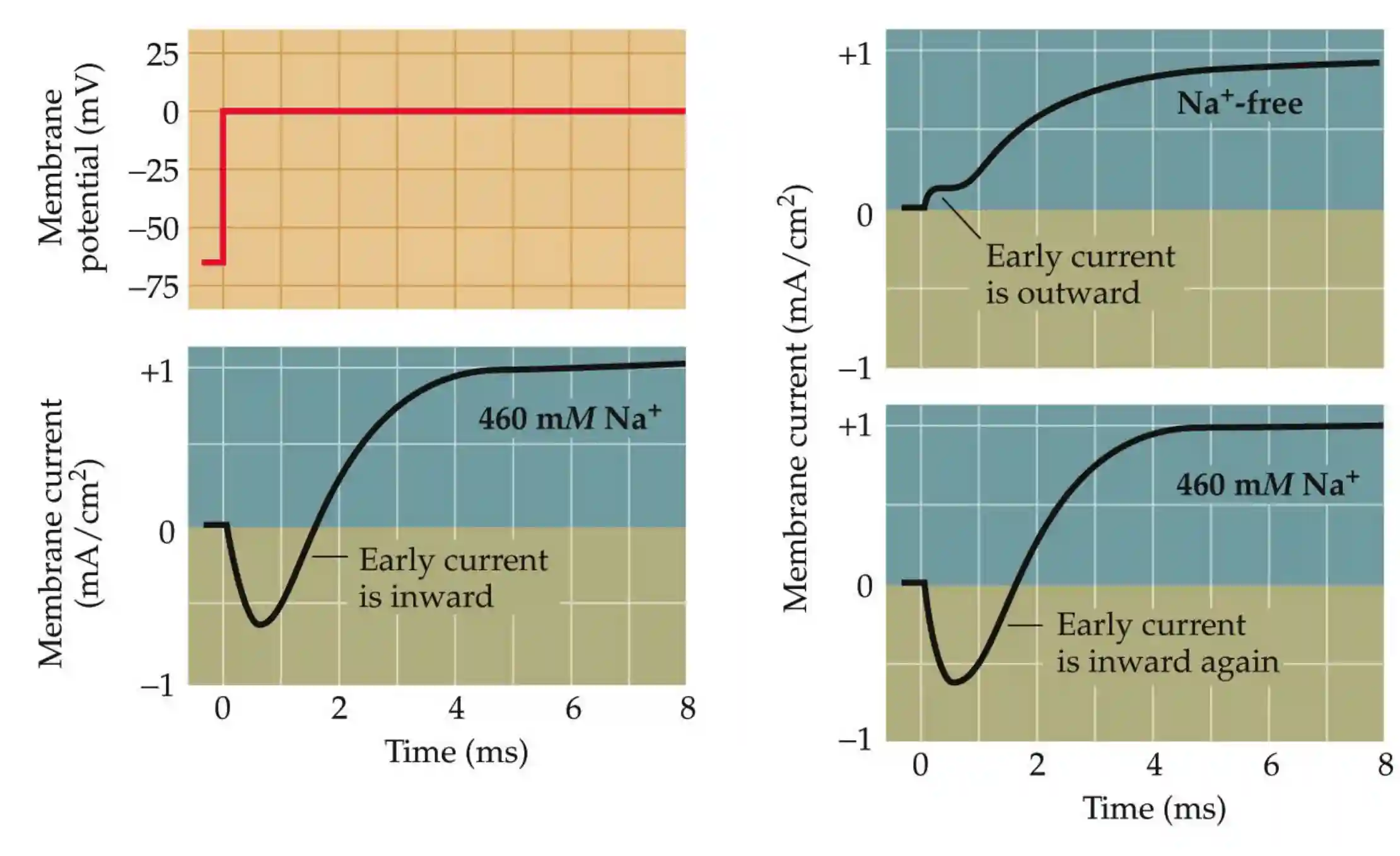 Sodium is important to have an early inward membrane current.
Then we have a potassium current outside and tetraetilammonium blocks that.
Sodium is important to have an early inward membrane current.
Then we have a potassium current outside and tetraetilammonium blocks that.
The conductance of those two follow a sigmoid with respect to membrane potential. The speed by which they activate is different. Potassium channels are a lot slower (about 1ms delay with respect with the Na channels).
Neurotoxins block action potentials
Depolarization and Hyperpolarization
- At rest, neurons are in a hyperpolarized state (negative membrane potential, typically around -70 mV).
- When a negative current is passed, the cell membrane just addapts passively and becomes a little bit less negative.
- When they receive input (e.g., touch, neurotransmitters), they become depolarized (more positive).
- If the depolarization reaches a threshold, called threshold potential, an action potential is triggered, and we see a spike. The number of spikes is related to the intensity of the action potential.

Example of a neuron with a threshold potential and hyper polarization
The Hodgkin-Huxley Model
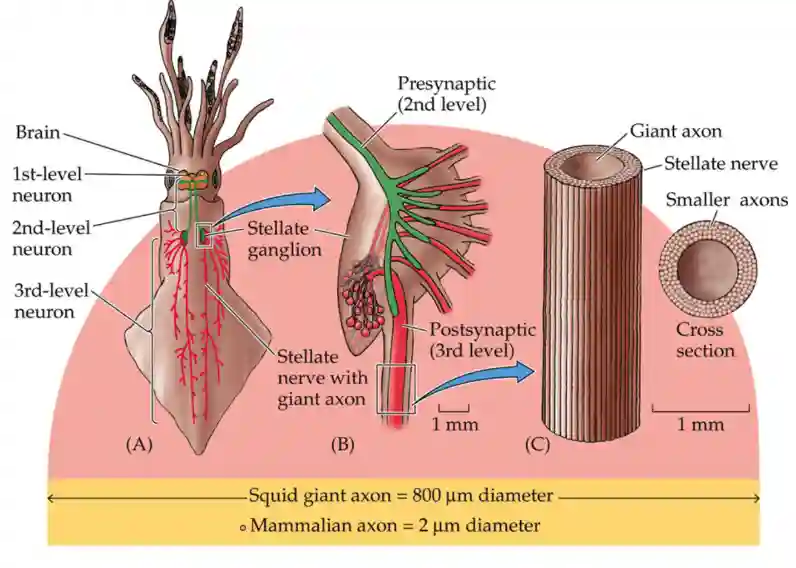
Alan Hodgkin and Andrew Huxley pioneered the study of action potentials in neurons using electrophysiology.
Experimental Setting
- They inserted electrodes into the axons of the giant squid (which has thick axons ~800 μm in diameter, ideal for measurements, hundreths of times larger compared to mammalian axons).
- They are larger for better conductivity, which helped them having faster reaction times.
- Measured how ion permeability changes over time during an action potential.
- Demonstrated that K⁺ is the dominant ion at rest, while Na⁺ permeability increases during an action potential, and returns with dominant K at rest (higher concentration in the cell than outside).
Key Findings
- Ion Concentration Dependence
- They determined that the action potential depends on Na⁺ and K⁺ gradients, but not Cl⁻.
- Increasing external K⁺ concentration disrupts the resting potential, making neurons more likely to fire spontaneously.
- Voltage-Dependent Ion Channels
- Na⁺ channels open at depolarization (threshold level ~ -55 mV).
- K⁺ channels open later and restore resting potential.
- Both channels show voltage-dependent gating:
- Na⁺ channels open rapidly, allowing inward Na⁺ current (depolarization).
- K⁺ channels open slower, allowing outward K⁺ current (repolarization).
- Logarithmic Graph of Potassium vs. Energy Level
- Hodgkin and Huxley plotted log K⁺ vs. voltage, showing a predictable relationship (consistent with the Nernst equation).
- Mathematical Modeling of Action Potentials
- They wrote a set of differential equations describing ion channel kinetics.
- These equations successfully predicted experimental results, marking the beginning of computational neuroscience.
- The equations describe:
- Membrane capacitance (how the membrane stores charge).
- Ion conductance (how easily ions pass through).
- Time-dependent changes in channel permeability.
- Sodium Permeability and Action Potentials
- Action potentials depend on a higher concentration of Na⁺ outside the cell.
- Hodgkin and Huxley confirmed that removing extracellular Na⁺ eliminates action potentials.
- Cl⁻ was shown to be unimportant for action potential generation.
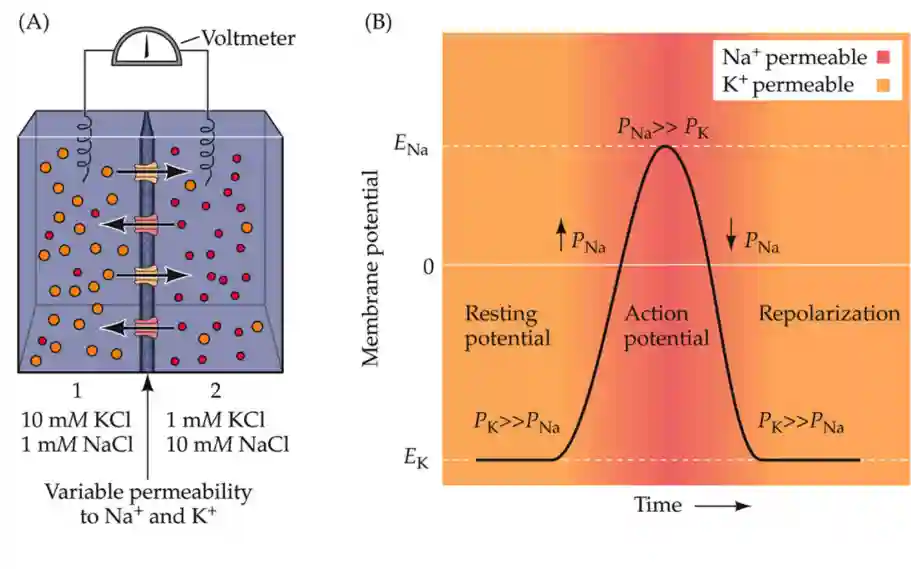
Resting Membrane Potential
Resting membrane potential mostly depends on the concentration of the ion in the extracellular space. The resting membrane potential depends mainly on the extracellular potassium concentration.
We break the electric gradient if there is too much potassium on the outside. On resting conditions potassium is the main variable.
![[Neural mechanisms-20250824151911795.webp| (a) The resting membrane potential depends mainly on the extracellular potassium concentration. (b) The measurements (red curve) deviate from the prediction of the Nernst Equation (black) at low outside potassium concentrations.]]
Role of Sodium in the Action potential
The shape of the membrane current depends upon sodium. With more sodium, the process is faster, the spike is a lot larger (they experimented with more sodium in extracellular environment). The above are the main conclusions.

(A,B,C): The membrane potential transiently becomes positive during an action potential due to the positive reversal potential of Na+ (D, E) The peak action potential amplitude (red) depends crucially on the extracellular Na+ concentration, but Na+ has little influence on the resting membrane potential. There is a deviation from the theoretical prediction (black) at low sodium concentrations.
Nernst Equation
Where is the gas constant, is the temperature, is the valence of the ion, is the Faraday constant, and and are the concentrations of outside and inside the cell, respectively. If we remove the constants and use log with base then the constant is . 10 fold change in concentration is about 58 volts of difference. So at equilibrium (outflux equals influx) we have 58 voltages. Assuming valence :
Where ION in this case is the potassium ion.
Examples of electrostatic equilibrium
To reinforce and extend the concept of electrochemical equilibrium, con- sider some additional experiments on the influence of ionic species and ionic permeability that could be performed on the simple model system in Figure 2.4. What would happen to the electrical potential across the membrane (the potential of side 1 relative to side 2) if the potassium on side 2 were replaced with 10 mM sodium (Na+) and the K+ in compartment 1 were replaced by 1 mM Na+? No potential would be generated, because no Na+ could flow across the membrane (which was defined as being permeable only to K+). However, if under these ionic conditions (10 times more Na+ in compartment the K+-permeable membrane were to be magically replaced by a mem- brane permeable only to Na+, a potential of +58 mV would be measured at equilibrium. If 10 mM calcium (Ca2+) were present in compartment 2 and 1 mM Ca2+ in compartment 1, and a Ca2+-selective membrane separated the two sides, what would happen to the membrane potential? A potential of +29 mV would develop, because the valence of calcium is +2. Finally, what would happen to the membrane potential if 10 mM Cl– were present in com- partment 1 and 1 mM Cl– were present in compartment 2, with the two sides separated by a Cl–-permeable membrane? Because the valence of this anion is –1, the potential would again be +58 mV.
There is a osmotic gradient, since the membrane is semipermeable only permeable to one kind of Ion (K). This is kept by the pumps. Osmotic force = electric force in the case of equilibrium.
The Voltage Clamp Technique
This was nobel prize winning technique to describe the voltage clamping technique. We have a method to keep the voltage of the axon to a fixed state and observe how the membrane potential changes, and we see that this potential changes with respect to the voltages.

Image from the slides
Technique by Kenneth Cole in the 1940s. This device is called a voltage clamp because it controls, or clamps, membrane potential (or voltage) at any level desired by the experimenter.
They discovered that the current produced by membrane depolarization changes in a manner dependent on the current.
It allows to:
- Measure the actual membrane potential using a microe-lectrode.
- Compare it to a command voltage that the experimenter wants to hold.
- Inject current through the electrode to keep .
- Record the injected current — by Ohm’s law, this is equal and opposite to the net ionic current flowing across the membrane:
Action potentials
Action Potential: A Rapid Electrical Signal
An action potential is a rapid, transient change in membrane voltage that propagates along the axon. It is fundamental to neural communication and muscle contraction.
Different action potential shapes
Why the action potential mechanism?
- Timing (precision)
- Generative aspect (avalanche of signals, not passive propagation but repropagation)
- It is fast?

Action potential shapes can vary across neurons: A) squid giant axon, B)myelinated axon, C) near cell body, D) inferior olive neuron, (E) Purkinje cell.
Amplitude and frequency
the amplitude of the action potential is independent of the magnitude of the current used to evoke it; that is, larger currents do not elicit larger action potentials. The action potentials of a givenneuron are therefore said to be all-or-none, because they occur fully or not at all
the intensity of a stimulus is encoded in the frequency of action potentials rather than in their amplitude. This arrangement differs dramatically from receptor potentials, whose amplitudes are graded in proportion to the magnitude of the sensory stimulus, or synaptic potentials, whose amplitude varies according to the number of synapses activated and the previous amount of synaptic activity.
Phases of the Action Potential
- Rising Phase (Depolarization)
- Voltage-gated Na⁺ channels open.
- Sodium influx makes the inside of the neuron more positive.
- Overshoot Phase
- Membrane potential becomes more positive than 0 mV.
- The inside of the neuron is now briefly positively charged compared to the outside.
- Falling Phase (Repolarization)
- Na⁺ channels inactivate.
- Voltage-gated K⁺ channels open, allowing potassium efflux, restoring a negative charge inside the neuron.
- Undershoot Phase (Hyperpolarization)
- K⁺ channels remain open too long, making the neuron even more negative than the resting potential.
- The neuron is temporarily less likely to fire another action potential.
The ReLU Activation Function
The Rectified Linear Unit (ReLU) activation function used in artificial neural networks is loosely inspired by biological action potentials.
- Threshold-like behavior: Like a neuron, ReLU only activates when input exceeds zero, similar to how neurons fire only when a threshold potential is reached.
- No negative output: Biological neurons do not fire negatively; they either remain inactive (hyperpolarized) or fire a positive action potential.
- Computational efficiency: ReLU simplifies calculations, much like how neurons minimize energy use by firing only when necessary.
However, real neurons exhibit more complex non-linearity, including:
- Adaptation (changes in firing patterns over time).
- Refractory periods (temporary inactivation after firing).
- Subthreshold activity (graded responses below threshold).
Thus, while ReLU is inspired by neuroscience, it oversimplifies real neural dynamics.
Generating an Action Potential
We can revise this model by adding parallel sections for , ions and leaked current. The current in the axon is a self-regenerating current trace, that allows to propagate it over long distances, with a certain speed. This is passively propagated, until the active component goes and regenerates the current.

Image from the book
We have refractoriness that prevents the action potential to activate again after a very short time frame.
Nodes of Ranvier
We talk about the saltatory condution in Firing-rate based Network models.
Nodes of Ranvier are gaps in the myelin sheath along the axon. It takes from the name of the discoverer of these nodes. Myelinated sections make the current flow 150x times faster compared to non Myelinated sections. It is believed that myelinated section originated from a virus that joined the cell, and now they are in symbiosys.
They have a lower threshold for triggering an action potential, and higher concentration of Na molecules. The distance between each section is about 1.5mm. This is made so that the decrease in potential is not so much, due to the distance (the potential halves every around 0.4mm with normal membrane values). The lower threshold helps to recreate the wave.
The voltage decay is lower in insulated versions, because the current is not lost to the environment (higher resistivity of the membrane thanks to the fatty part). The myelination makes thin nerve fibers even faster than big squid axons! But they are quite expensive to maintain and build.
The propagation of voltage change within an insulator (the myelin) is fast, while the normal unmyelinated propagation is usually slower.
For example the large squid axon has a speed of 20m/s, while fibers of myelinated fybers have 100m/s speed.
Squid Axon
Why some neurons are unmyalinated
- Some neurons just don't need it because they are short connections.
- Some neurons benefit from slow connections.
- So that we have some finetuning in signal speed across different sources. (e.g. some retina neurons)
- It is costly to maintain and produce the myelinated fibers.
Squid vs Mammalian Neurons Environment
| Ion | Intracellular (mM) | Extracellular (mM) |
|---|---|---|
| Squid neuron | ||
| Potassium (K⁺) | 400 | 20 |
| Sodium (Na⁺) | 50 | 440 |
| Chloride (Cl⁻) | 40–150 | 560 |
| Calcium (Ca²⁺) | 0.0001 | 10 |
| Mammalian neuron | ||
| Potassium (K⁺) | 140 | 5 |
| Sodium (Na⁺) | 5–15 | 145 |
| Chloride (Cl⁻) | 4–30 | 110 |
| Calcium (Ca²⁺) | 0.0001 | 1–2 |
Generating and Action Potential
Neurons communicate via electrical signals generated by the movement of ions across the cell membrane. These movements are mediated by two primary types of membrane proteins:
Ion Transporters vs. Ion Channels
- Ion Transporters (Active Transporters)
- Move ions against their concentration gradient using ATP.
- Example: Na⁺/K⁺ ATPase (Sodium-Potassium Pump), which pumps 3 Na⁺ out and 2 K⁺ in, maintaining a concentration gradient.
- Establish the chemical and electrical gradients necessary for neuron function.
- Ion Channels
- Allow ions to diffuse down their concentration gradient (passive transport).
- Highly selective for specific ions (e.g., K⁺ channels, Na⁺ channels).
- Can be voltage-gated (open/close based on membrane potential) or leak channels (always open).
- The osmotic gradient drives ions, but the membrane potential balances the flow.
Chloride is used to hyperpolarize the cells.
Speed of membrane potential change
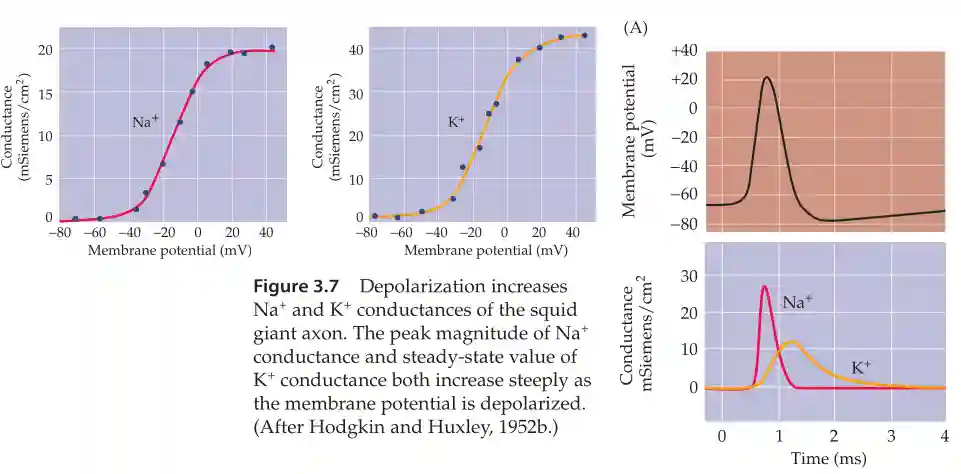
This is why the action potential is about 1ms (which makes it faster compared to longer values). Basically, the sodium ion channel opens and closes very quickly. So:
- The sodium channels are faster than potassium channels
- Sodium goes in, potassium goes out.
- Potassium going out makes it balanced.
- Sodium going in makes it depolarized.