Vision is THE most important sense for humans. Most of the information we get is through vision
- 90% vision
This is true for humans, but for mice for example it is different, they have probably a 64x64 pixel resolution equivalent. For humans, visual data is more important, it is faster compared to speech and other senses.
We can also see this when we are talking about quantity of neurons:
- 30% of neurons in the cortex are for vision.
- 8% tactile, touch
- 3% hearing
- Rest is for other
Precision of human vision
Human vision is estimated to be about 576 Megapixels of data (3M snapshots patched together with saccades, that has that pixel image value), since it can distinguish 0.6arc-minutes (0.01 degrees). There is an estimate of about 60kk ± 12kk rods and 3kk ±0.5kk cones. This means that the image is constructed by high resolution saccades (patches).
it appears to be best suited to extract the sort of information that may be useful to animals, including humans, in a natural environment. (Squire et al. 2012)
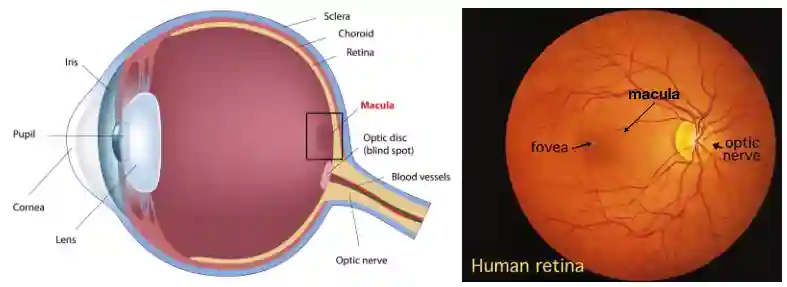
Eye Anatomy
We can change the shape of the lens to focus on objects of different distance.
Saccades
To get the full view of the environment humans are used to do saccades. Brain directs it to look for the spaces that are important, the key features. Monitoring this is important for parts like (Leslie 2001).
Receptive field
We have talked about this in Neural mechanisms, Convolutional Neural Network but also in Autoregressive Modelling. So you should know this topic.
The Retina
Basic Anatomy
The retina is able to reshape its lens, to focus on close or far objects. This part is transparent, there are some cells that register the information and send it in the optical nerve. There is one spot in the retina that has no receptors, it is called blind spots. Density of receptors define the resolution of the image you get.
The Macula contains many Cones, usually good for color detection, it is important for sharp vision. The fovea is the center of the macula, it has very very high concentration of cone cells, exclusively those.
Neurons in the Retina
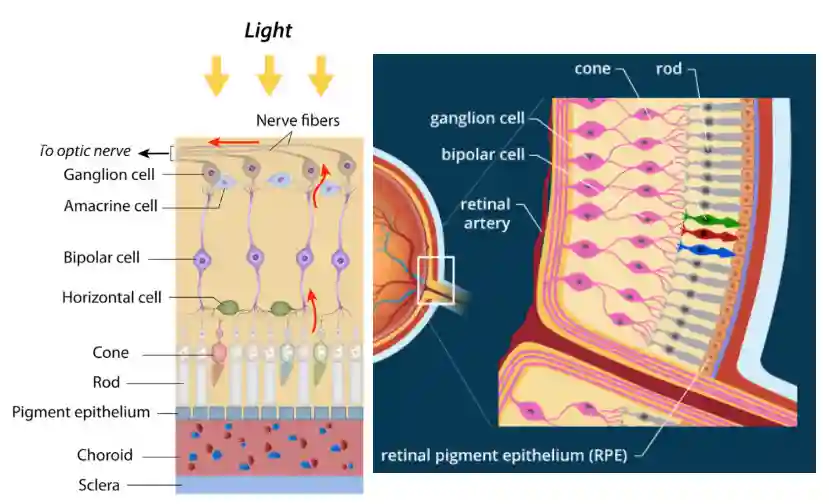
Bipolar cell and horizontal cells already do process information. Only the ganglion cell is spiky, the others are analogic, since it has better precision.
Horizontal cells are inhibitory (GABAergic) and, as the name implies, they contact photoreceptors over a much larger horizontal extent of retina than bipolar cells. Thus, because they are the only neurons that sample over a sufficiently large area in the outer plexiform layer, horizontal cells are thought to be responsible for the antagonistic surround seen in the receptive fields of bipolar cells. (Squire et al. 2012).
Horizontal Cell
- Lateral Inhibition When one photoreceptor is strongly activated by light, horizontal cells inhibit neighboring photoreceptors, which enhances contrast and edge detection. So they help to spread the information in this manner.
- Feedback to photoreceptors:
- They provide negative feedback to photoreceptors, helping to adjust gain depending on the overall light level.
- This contributes to phenomena like center-surround receptive fields in bipolar and ganglion cells.
- Color processing:
- Horizontal cells can connect to cones of different types (e.g., S, M, L cones) and help with opponent color processing.
Spiking Ganglion Cells
Ganglion Cells are the output of the retina, it is where the information is sent to the brain, it takes about 10ms to process this information and exit the eye.
Other cells do not spike, they continuously send the signal. Ganglion cells are the only cells that spike here (digital signals have long range without decay (see Neural mechanisms, while analog signals have this problem, for short distances they are great). They have tenths of centimeters to travel before reaching the objective.
Photoreceptor Layer
In the photoreceptor layer, Cones and rods live. Their role is to capture the light, and process it locally before sending it to the brain.
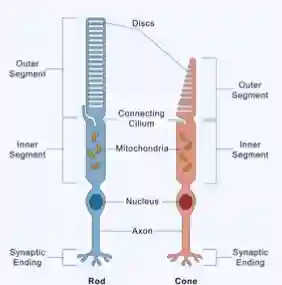
Rods and Cones
- Rods: Sensitive to low light (scotopic vision); no color perception.
- Cones: Active in bright light (photopic vision); responsible for color and detail.
Rods
- Shape: Long, cylindrical outer segment.
- Outer segment: Contains many stacked membranous discs loaded with rhodopsin (photopigment).
- Size of outer segment: Longer and thinner than cones, allowing more photopigment → highly sensitive to light.
- Synaptic terminal: Large, single connection (spherule) with bipolar cells.
- Location in retina: More numerous in the peripheral retina.
Cones
- Shape: Shorter, tapered outer segment (cone-shaped).
- Outer segment: Fewer, continuous membranous folds with cone opsins.
- Size of outer segment: Shorter, thicker → less sensitive to dim light but allows color discrimination.
- Synaptic terminal: Pedicle with multiple connections to bipolar and horizontal cells.
- Location in retina: Concentrated in the fovea (central retina) for high-acuity vision.
These rods have synapses, but they are very special. The optic disks contain opsins, which interact to light and are able to communicate this signal to the other parts.
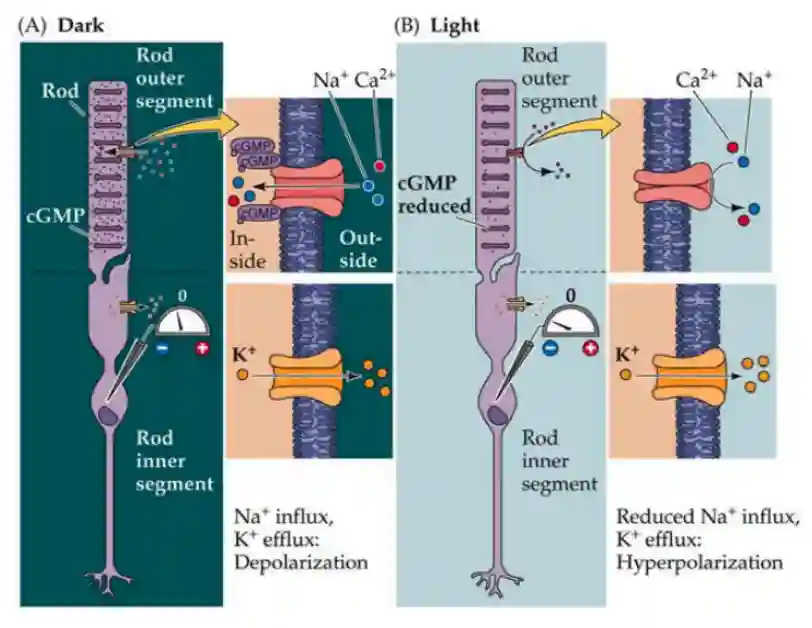
Light Sensitivity Processes
We will explain the process of how photoreceptors are hyperpolarized by light.
In the disks of the Rods and Cones there are opsins and rhodopsins, proteins that react to light.
- Activates transduction of g protein transduction (it is released).
- This hydrolizes cGMP to GMP, activating an ion channel for sodium, which is now closed.
- This changes the membrane potential of the rod or cone.
Opsins are S, M, L for blue green and red light. Rhodopsins are for rods.
In the dark, the channels are open, so you have influx and pumps for potassium, getting depolarized. In light, you are pushing out potassium, so the cell is getting more negative in membrane potential -> hyperpolarized.
In this way, light increments lead to hyperpolarization and a reduction in neurotransmitter release, whereas light decrements lead to depolarization and an increase in transmitter release. (Squire et al. 2012)
The cones and rods are also specialized in some forms of wavelengths. Cones have different lengths to get different wavelengths (three forms Short, Medium and Long), different humans have different sensitivity. This difference is also why humans and animals are sensitive to different wavelengths.
There has been some research in genetic engineering cross animals and nano-antennaes to convert infrared to visible light.
Recycling the Retinol
Rhodopsins change conformation when activated and need to be recycled.
In the disk, the cis-isomer is transformed into a trans-isomer, which are not usable anymore for the light, these are then recycled using some enzymes. Recycled in the retinal pigment epithelium. There is some transport to have it in the outer segment.
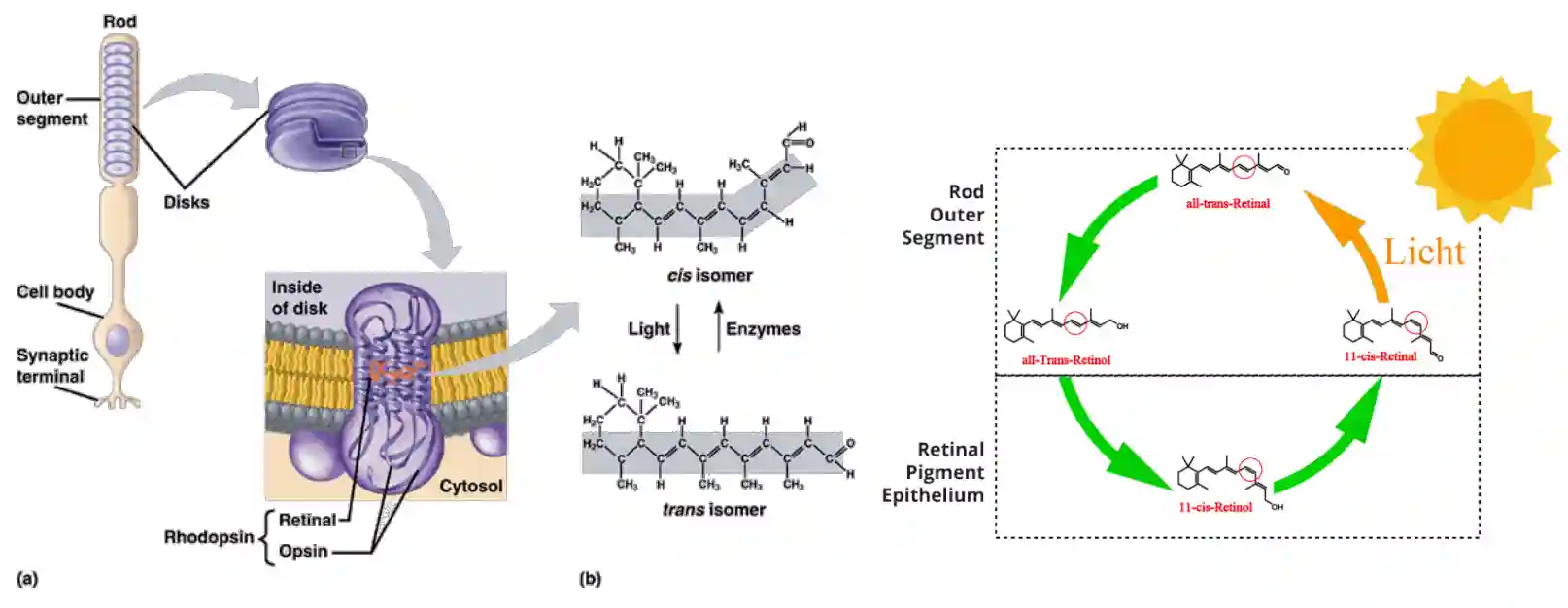
The 11-cis-retinal is a form of Vitamin A.
Differential responses of Rods and Cones
Under scotopic conditions when luminance levels are very low (e.g., starlight), only the rods are active. As luminance levels increase to mesopic conditions (e.g., moonlight), both the rods and cones contribute to vision. As luminance levels increase further yet, to scotopic condition (e.g., sunlight), rod responses saturate and only the cones contribute to vision. (Squire et al. 2012)
Rods -> scotopic vision, concentrated for black and white vision Cones -> photopic vision, concentrated for color and detail (you need a lot of light to activate cones).

Rods have many many rods connected to the rod bipolar cell, while cone is single and focused.
! Showing rod and cones’ interplay with different light conditions.
{kind=link}
Retinal Epithelium Recycling cycle
When new disks are formed, old disks accumulate at the top of the rod, they get engulfed in pigment epithelium and separates from the rod.

The experiment in the image (right) shows the idea of disk replacement. Some radio amminoacids are used to build the disks, they move up and end up in the pigment epitelial cell
Rods and cones are shed following the circadian rhythm.
Mainly used to recycle the components and remove the waste, produce the 11-cis-retinal molecule. Prevents disfunctional discs.
The disks provide several benefits:
- Light absorption -> Photoactivation: Photon is absorbed → 11-cis-retinal isomerizes to all-trans-retinal → rhodopsin becomes active (meta II) and activates transducin → phototransduction cascade.
- Growth factors
- Phagocitosis?
- Visual Cycle
- And metabolism. I have no idea how.
Distribution of Photoreceptors
Color only where you focus, since it is more complex, in the fovea mostly we have all cones.
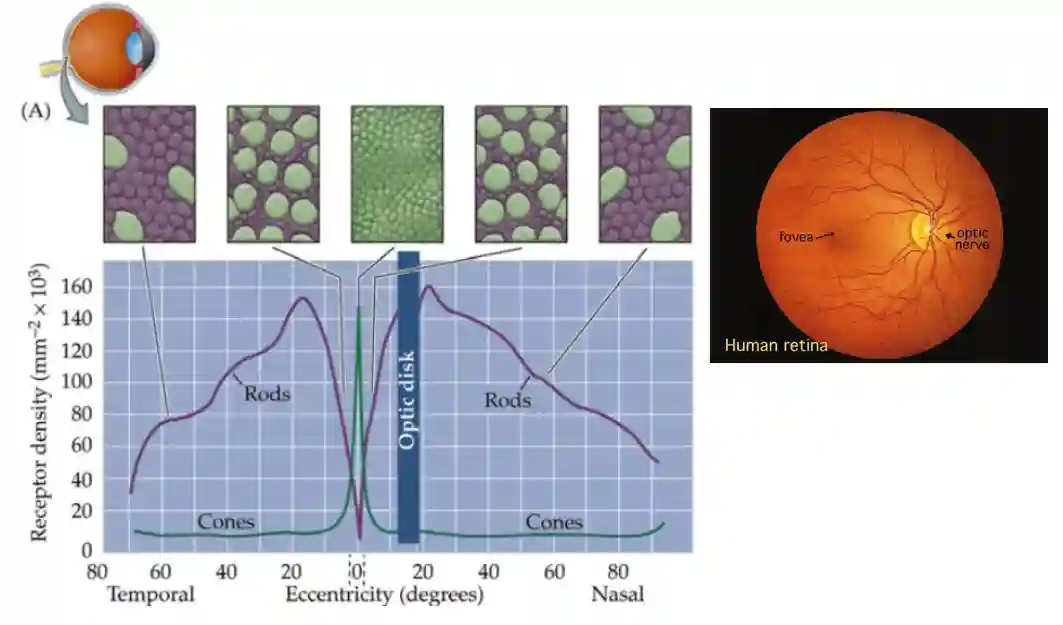
! Distribution of color sensitivity for Cones, and absorbance of rods too.
{kind=link}
Different animals have different expression of rods and cones (and different opsin expressions, that allow for different light sensitivity). With genetic editing, it is possible to make it available for some animals too.
On and Off center cells
Bipolar cells behave in opposite ways. They are connected to separate pathways of ganglion cells.
Increase and decrease interleaved patterns, they flip their outputs, quite easily.
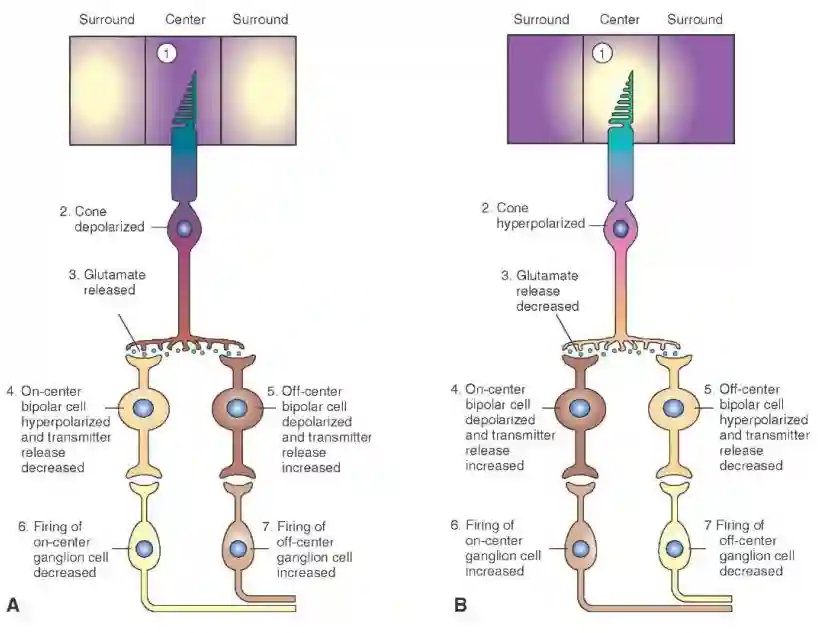
If you have surround, then they both spike, but a little bit less. You can track if a ganglion cell is on center or off center. This is probably useful to detect movements in the sides, probably good to detect edges of the image, useful for the V1 pathways in vision, see Architecture of the Brain, see paper from olshausen and Field, (Olshausen & Field 1996).
There is an experiment of Kuffler 1951 which is close to this. Hubel and Wiesel. (Hubel & Wiesel 1962).
Pathways
The signals gets into Superior colliculus, that handles saccades and eye movements. Gets also to the lateral geniculate nucleus that then projects to the V1 (to check). The pretectum has reflex for pupil and lenses. See Architecture of the Brain.
Retinal Visual Projections
Part of the image are not available to both eyes, they got to part of the optic tracts.
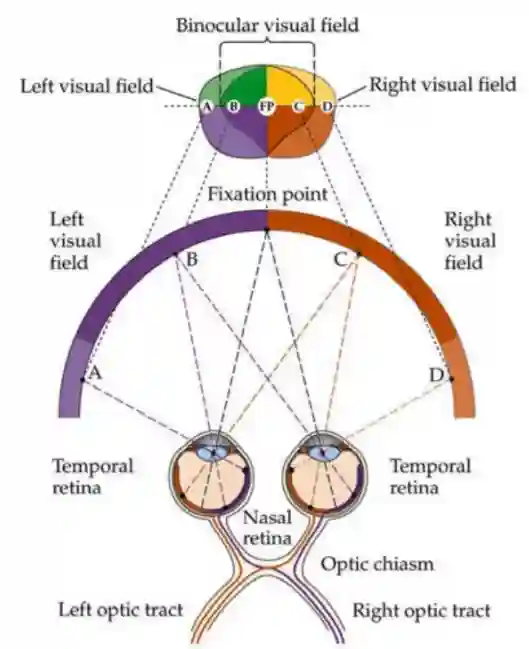
Projections to the Visual Cortices
The processing in the visual cortex is flipped but this is ok.
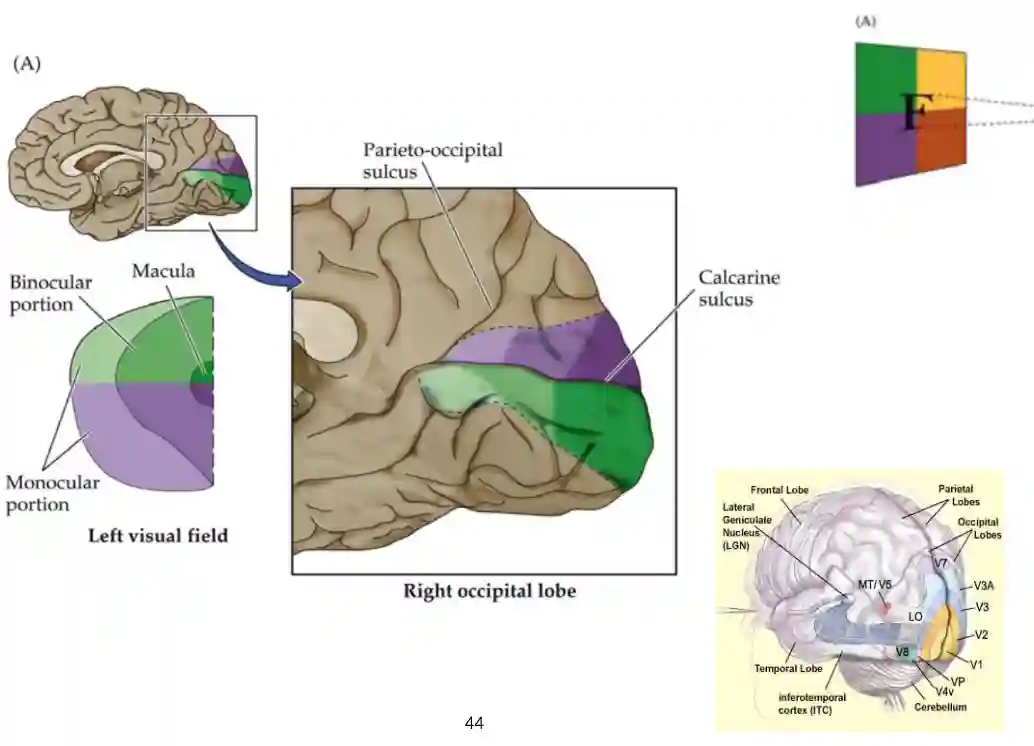
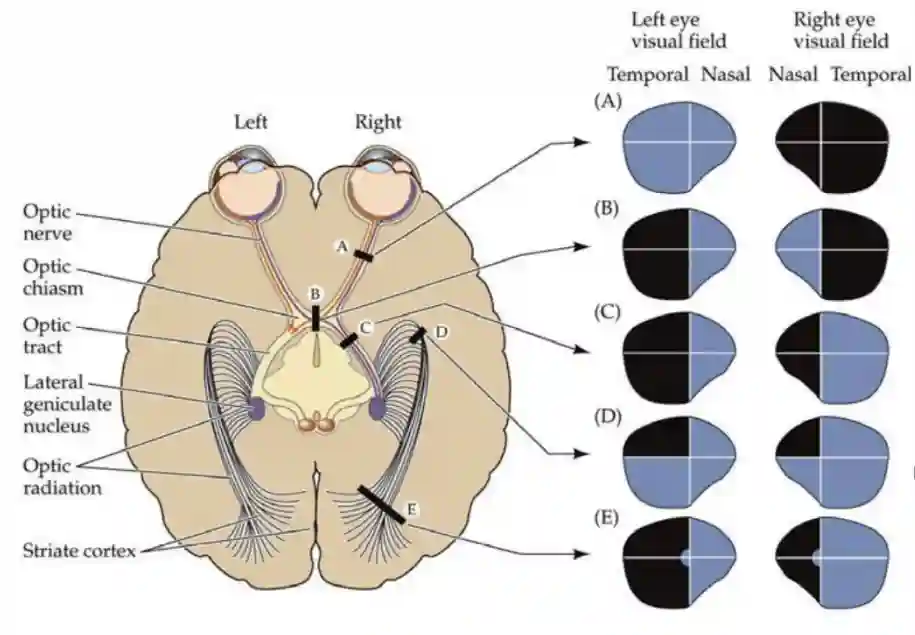
Visual Pathways
This is the standard pathway:
- Retina → Optic nerve → Optic chiasm → Optic tract
- Most fibers go to the lateral geniculate nucleus (LGN) of the thalamus.
- Superior colliculus has feedback, and gives eye movement directions.
Retina
├─ LGN → V1 → Dorsal stream ("where/how")
├─ LGN → V1 → Ventral stream ("what")
├─ Superior colliculus (eye movements)
├─ SCN (biological clock)
└─ Pretectal nuclei (pupil reflex)
References
[1] Hubel & Wiesel “Receptive Fields, Binocular Interaction and Functional Architecture in the Cat's Visual Cortex” The Journal of Physiology Vol. 160(1), pp. 106--154 1962
[2] Squire et al. “Fundamental Neuroscience” Academic Press 2012
[3] Leslie “Theory of Mind” 2001
[4] Olshausen & Field “Emergence of Simple-Cell Receptive Field Properties by Learning a Sparse Code for Natural Images” Nature Vol. 381(6583), pp. 607--609 1996